Esta dirección de correo electrónico está protegida contra spambots. Necesita activar JavaScript para visualizarla.
El parasitismo es una estrategia ampliamente distribuida dentro del árbol de la vida, y las plantas con flor no son una excepción.
Las plantas parásitas tienen la habilidad de alimentarse directamente de otras plantas. Para ello, invaden los tallos o raíces de plantas hospedadoras mediante raíces modificadas llamadas haustorios (Figura 1). Éstas les permiten conectar su tejido vascular con el de la planta hospedadora posibilitando el intercambio de nutrientes, agua e incluso moléculas como ARN y ADN.
 |
| Figura 1: foto de un haustorio. Planta parásita Cassytha. |
Dentro de las plantas parásitas se distinguen dos categorías principales: hemiparásitas y holoparásitas (Figura 2). Las primeras se caracterizan por ser plantas capaces de fotosintetizar, que absorben principalmente agua y solutos de sus hospedadoras. Por su parte, las holoparásitas comprenden el 10% de las plantas parásitas y representan la condición más extrema del parasitismo, ya que no tienen clorofila y son incapaces de realizar fotosíntesis, dependiendo completamente de su hospedador para vivir.
La transición desde el estilo de vida libre a la vida parásita, habría ocurrido de forma independiente en 12 linajes de plantas. Esta transición significó un cambio radical en la estrategia de vida de estos grupos, y en consecuencia estuvo acompañada de grandes modificaciones morfológicas y moleculares. En cuanto a la morfología, las hojas se redujeron hasta formar pequeñas escamas y las raíces se transformaron para reconocer e invadir a la planta hospedadora. A nivel celular y molecular, el cloroplasto sufrió reducción de su genoma, degradación de membranas fotosintéticas, pérdida de clorofila y capacidad de fotosintetizar, en algunos casos.
Otro mecanismo que participa en la evolución de estas plantas es la adquisición de material genético proveniente de especies no emparentadas: la transferencia génica horizontal (TGH). En los últimos años se han detectado numerosos eventos de transferencia de genes desde las plantas hospedadoras a las plantas que las parasitan. Esto sugiere que el parasitismo podría facilitar la TGH, aunque el alcance y las repercusiones de este fenómeno en las plantas parásitas todavía no están claros.
 |
| Figura 2: fotos de plantas hemiparásitas y holoparásitas. Fuente: Henning S. Heide-Jørgensen (2008) |
Transferencia génica horizontal
La transferencia horizontal es un proceso común en organismos procariotas (organismos sin núcleo) que intercambian ADN entre distintas especies. Sin embargo, el descubrimiento de eventos de TGH entre eucariotas (organismos con núcleo) es relativamente reciente. Entre los eucariotas, las plantas han sido reconocidas como dadoras y receptoras de material genético con diversos organismos; pero la transferencia planta-a-planta es la más frecuente, siendo mucho más común en las mitocondrias que en los cloroplastos o en el núcleo.
La transferencia de material genético entre especies no emparentadas requiere que éstas entren en contacto. Se han propuesto distintos mecanismos de contacto entre plantas, algunos de forma directa, como pueden ser la íntima relación parásito-hospedador y la formación de injertos naturales, o de forma indirecta, a través de vectores biológicos, como granos de polen (polinización ilegítima), virus, bacterias, hongos e insectos.
En el caso particular de las plantas parásitas, se ha propuesto que mitocondrias completas de la planta hospedadora atraviesan el punto de contacto e ingresan a las células del parásito. Dado que las mitocondrias, a diferencia de los cloroplastos, se fusionan frecuentemente in vivo, las mitocondrias de ambas plantas se fusionarían y sus genomas podrían interactuar e intercambiar fragmentos de ADN, dando lugar a genomas mitocondriales quiméricos que contienen genes foráneos (genes de la otra planta) (Figura 3). Esto a su vez explicaría por qué no se observa ADN foráneo en los cloroplastos.
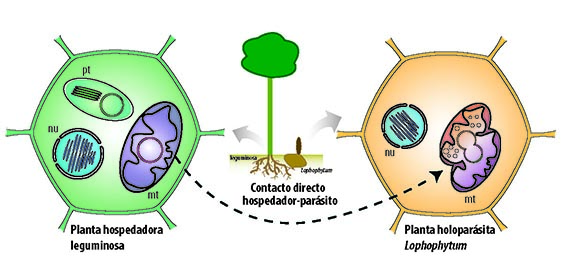 |
| Figura 3: Modelo de fusión de mitocondrias |
Pero, si esta transferencia ocurre en una única célula… ¿cómo se explica que todas las células de un organismo posean genes foráneos? Esto sucede gracias al carácter totipotencial de las células vegetales (es decir que cada célula puede generar por multiplicación cualquier estructura u órgano), la célula con mitocondrias quiméricas podría formar estructuras reproductivas que resultarían en óvulos, y por lo tanto en embriones, poseedores de mitocondrias quiméricas.
Plantas holoparásitas del género Lophophytum
El Grupo de Genómica Evolutiva de Plantas, liderado por la profesora M. Virginia Sánchez Puerta de la Facultad de Ciencias Agrarias de la UNCUYO, se ha centrado en el estudio de la evolución molecular de plantas holoparásitas del género Lophophytum (familia Balanophoraceae) con el fin de comprender la dinámica, evolución, e incidencia de la TGH. Lophophytum es un género exclusivamente sudamericano y comprende cinco especies, de las cuales dos se encuentran en la Argentina: Lophophytum mirabile en el noroeste, y Lophophytum leandri en el noreste.
Estas especies parasitan a plantas leguminosas de la tribu Mimosoideae (familia Fabaceae): Anadenanthera colubrina y Parapiptadenia rigida, respectivamente (Figura 4). Estas parásitas se conectan a las raíces de su hospedador y desarrollan un cuerpo vegetativo o tuber totalmente subterráneo, del cual sólo emergen a la superficie las inflorescencias (Figura 4).
La docente Sanchez-Puerta, junto con estudiantes de grado y posgrado, se embarcaron en la secuenciación y caracterización del genoma mitocondrial completo de Lophophytum mirabile. Este estudio permitió conocer la estructura del genoma, su contenido y la forma en la que está organizado. Análisis evolutivos de cada uno de los genes presentes en el genoma mitocondrial revelaron la frecuencia (i.e. número de secuencias transferidas), el origen (i.e. quien fue su dador), y el impacto de la TGH en el genoma mitocondrial de la parásita.
Un genoma mitocondrial sorprendente
El genoma de la mitocondria de Lophophytum mirabile tiene un tamaño de 800 mil pares de bases, y está organizado en 54 cromosomas circulares, que poseen tan sólo 55 genes. Más del 75% de dichos genes fueron obtenidos de su hospedador por transferencia horizontal. Igual de impactante es el hecho que 26 de los 54 cromosomas no poseen genes y su función es desconocida como así también la razón por la cuál son conservados por la planta parásita.
 |
| Figura 4: Foto de Lophophytum mirabile y su hospedador Anadenanthera colubrina |
Impacto evolutivo de la adquisición de genes foráneos
Una de las preguntas más importantes en el campo de la biología evolutiva es si los genes foráneos son funcionales y, si lo fueran, ¿cuál sería su impacto evolutivo? En la mayoría de las plantas los genes foráneos no son funcionales y parecen tener muy bajo efecto en la evolución de las mismas. Sin embargo, en el caso de Lophophytum mirabile, los genes foráneos parecen estar activos y habrían reemplazado las funciones de los genes originales que se perdieron en la evolución de esta planta.
Los genes adquiridos por la holoparásita estarían a cargo de la producción de energía a través de la respiración. Este descubrimiento es muy novedoso, ya que hasta el momento se creía que la transferencia horizontal de genes no cumplía ningún rol de importancia en la vida y evolución de las plantas. Los resultados de este estudio despiertan nuevos interrogantes sobre el impacto de la TGH en la relación parásito-hospedador y en la co-evolución de los genomas nucleares y mitocondriales de las plantas.
¿Una planta sin cloroplastos?
Otro descubrimiento sorprendente es la ausencia de cloroplastos en Lophophytum. Si bien ya había sido documentada la pérdida de fotosíntesis en muchas plantas holoparásitas, los cloroplastos cumplen también otras funciones celulares y han sido conservados aún en ausencia de clorofila. El estudio genómico de Lophophytum mirabile parece indicar que el cloroplasto y su genoma plastidial han sido eliminados durante la evolución de esta holoparásita.
Referencias
Barkman, T. J., et al. (2007). Mitochondrial DNA suggests at least 11 origins of parasitism in angiosperms and reveals genomic chimerism in parasitic plants. BMC Evol. Biol., 7 (1), 248.
Ceriotti, L.F, Wohlfeiler, J & MV Sanchez-Puerta. (2014) Estudio evolutivo de plantas holoparásitas del género Lophophytum (Balanophoraceae). JBAG S25: 248.
Gonzalez, A.M. and Mauseth, J.D. (2010). Morfogenesis is highly aberrant in the vegetative body of the holoparasite Lophophytum leandrii (Balanophoraceae): all typical vegetative organs are absent and many tissues are highly modified. Int.J.PlantSci.171 (5): 499–508.
Heide-Jorgensen HS (2008). Parasitic flowering plants. Leiden: Koninklijke Brill NV.
Mower, J.P., et al. (2004). Plant genetics: gene transfer from parasitic to host plants. Nature, 432, 165-66.
Sato, H.A. y Gonzalez, A.M. (2013). Anatomía y desarrollo de la flor estaminada, microsporogénesis y microgametogénesis en especies de Lophophytum (Balanophoraceae) en la Argentina. Bol. Soc. Argent. Bot. 48 (1): 59-72.
Westwood, J.H., et al. (2010). The evolution of parasitism in plants. Trends Plant Sci., 15, 227-35.